深圳华大生命科学研究院联合华中农业大学等团队,利用华大自主研发的Stereo-seq时空组学技术,结合多切片策略,在单细胞分辨率水平上,对斑马鱼受精后24h(hpf, hour post-fertilization)以内6个关键时间点(3.3、5.25、10、12、18、24hpf)的胚胎进行研究。研究人员首次绘制了斑马鱼胚胎早期发育的时空转录组图谱,构建了在组织空间维度上的发育轨迹。此研究能够直观地观察到,在原位组织上细胞是如何分化、迁移并发育形成各组织器官的。该文章于2022年5月4日发表在Developmental Cell,以下是文章第一作者刘畅博士的详细解读。

文章题目:Spatiotemporal mapping of gene expression landscapes and developmental trajectories during zebrafish embryogenesis
发表时间:2022年5月4日
发表期刊:DevelopmentalCell
主要研究团队:深圳华大生命科学研究院、华中农业大学团队等
影响因子:12.270
DOI:10.1016/j.devcel.2022.04.009
研究背景
在严格的时空调控下,脊椎动物胚胎在较短时间窗口内,通过基因表达和细胞状态的剧烈改变促成发育,这是一个复杂的动态过程。其中,转录因子、信号通路、细胞外基质等多种因素在细胞增殖分化和细胞命运决定过程中发挥着重要的调控作用。在时间和空间纬度上,探究各个因素之间的分布关系和复杂调控,是解析胚胎发育的重要途径。
过去几年,利用单细胞组学技术,研究人员已经系统地揭示了斑马鱼如何从一个受精卵发育成完整个体的单细胞分子调控机制。然而,空间维度上理解转录组调控发育的机制,仍是全面认识发育的主要挑战。斑马鱼胚胎尺寸只有毫米数量级,目前,鲜有空间转录组技术能够满足其空间分辨率上的研究需求。
研究样本
空间转录组/单细胞转录组样本:受精后3.3、5.25、10、12、18、24h的斑马鱼胚胎。
研究策略
选取受精后24h内6个关键发育时间点的斑马鱼胚胎,对总共91张矢状面冰冻切片进行了Stereo-seq分析,搭建了斑马鱼早期胚胎发生时空转录组图谱在线开放资源库ZESTA (https://db.cngb.org/stomics/zesta/)。然后,利用基于计算基因表达空间分布相似性的空间共变基因模块,探究了空间功能区域间的相互调控作用,并对每个关键发育时间点的Stereo-seq和scRNA-seq数据进行整合,构建了斑马鱼胚胎发育过程中细胞命运转变和细胞分子变化的时空发育轨迹,探索性地研究了细胞空间微环境与分化方向之间的关联。最后,分析了斑马鱼胚胎发生过程中不同潜在互作配体-受体对的动态空间分布,发现了新的配体-受体对互作模式,提示了潜在的胚胎发育调控机制。
研究结果
1. 应用Stereo-seq技术获取斑马鱼早期胚胎发育的时空转录组数据
选取6个发育关键时间点(3.3、5.25、10、12、18、24hpf)的斑马鱼早期胚胎,分别制备成15μm厚度的冰冻切片,并把组织切片贴在时空芯片上进行Stereo-seq测序(图1A)。由于胚胎直径约为1mm,而芯片大小为1cm×1 cm,所以可以实现一张芯片上贴多张切片的策略。以24hpf胚胎为例,在2张芯片上对总共17张胚胎切片进行捕获测序,发现24hpf胚胎不同切片捕获到的UMI数和基因数相当(图1B、图1C),说明多切片策略能较好地消除不同生物样本之间的批次效应。同时,通过比较分辨率从143μm(bin200, 200*200 DNB)到10μm(bin15, 200*200 DNB,相当于1个细胞直径)的空间聚类效果,证实了在10μm分辨率下能够得到更加精细且准确的分群(图1D)。每个时间点的spots数量、UMI数以及基因数都足以支持后续数据分析(图1E~图1G)。因此,高分辨率的Stereo-seq时空组学技术,结合多切片策略,使研究人员对如斑马鱼胚胎这样的小尺寸生物样本进行发育图谱绘制成为了可能。在此,研究人员构建了一个高质量的斑马鱼胚胎发生时空转录组图谱(zebrafish embryogenesis spatiotemporal transcriptomic atlas,ZESTA)。

图1 对发育进程中的斑马鱼胚胎进行多切片、高分辨时空组Stereo-seq测序
2. 斑马鱼胚胎发育时空转录组图谱的空间聚类和分子特征
在10μm的分辨率下,研究人员将6个关键时间点的所有胚胎切片合并在一起进行无监督分群,得到了较为清晰的组织结构和空间特异性(图2A)。得益于Stereo-seq较高的空间分辨率,研究人员能够对不同的组织器官进行更为精细的细致分群,同时结合各个区域的marker基因,对空间位置相近的组织区域进行更加明确地划分。以3.3 hpf的胚胎为例,聚类能够得到3个具有明显空间分布差异的细胞分群:margin alblastomere、superficial blastomere、deep blastomere(图2B~图2E)。3个分群marker基因的空间分布、分化前后关系都与前人报道一致。此外,研究人员还发现了一些分群中未被报道的marker基因空间分布和原位杂交分布一致,表明了空间转录组数据的可靠性和新marker基因的发掘潜力。
斑马鱼胚胎在受精后发育到24h时,大脑的各个脑区已经有了明显的分界线,此时的神经系统已经发育得较为成熟。因此,研究人员针对24 hpf胚胎的神经系统进行了细致分群,得到了神经系统精密的空间转录组图谱,在超高分辨率条件下,进一步利用数据去揭示一些单细胞数据无法回答的问题(图2F、图2G)。例如,在后脑结构中,通过细致分群发现了一群位于后脑背侧的神经干细胞和位于后脑腹侧的神经前体细胞。对这两群细胞进行拟时序分析,得到的干细胞分支和神经前体细胞分支分别处于拟时序的先后位置(图2H)。根据这2个分支,细胞能够被进一步分成5个state,通过比较这些state细胞类群的差异表达,也能区分这些分支细胞类群的分化特性(图2I~图2K)。如state 1的细胞主要表达干性marker基因,而state 2这一分支的细胞高表达mRNA翻译相关基因,提示这一分支干细胞较state 1更趋向于分化,state 3的这群神经元前体细胞则更趋向表达神经元前体相关marker基因。同时,进一步结合每个state所处的空间位置,研究人员发现,state 1这群干细胞更倾向于分布在斑马鱼前侧和背侧,而state 2这群干细胞倾向于分布在斑马鱼后侧和腹侧。这一发现揭示了后脑神经干细胞的分化异质性空间分布,是对前人认知的一个补充。

图2 斑马鱼胚胎发育时空转录组图谱的空间聚类和分子特征
3. 空间共变基因模块揭示了斑马鱼胚胎不同功能区域之间的相互作用
基因在空间分布上存在差异,表达分布相似的基因所聚集形成的模块能够提示胚胎不同的功能区域。为了识别显示相似空间分布的共同变化基因,研究人员利用Hotspot计算基因表达的空间分布相似性,在每个胚胎上划分空间表达分布相似的共变基因模块。通过GO分析对每个模块进行注释,发现这些模块的功能与所处的空间位置以及所聚集的细胞类型都是高度一致的,提示Hotspot计算的准确性(图3A、图3C)。进一步围绕空间这一要素对斑马鱼胚胎发育进行挖掘,以便能够对空间共变基因模块之间的互作关系进行分析。如通过对其中分布范围广且包含多个细胞类型的中脑-后脑边界神经脊椎骨(M12)模块进行GO分析(图3B、图3D),发现与M12相关的生物功能与神经嵴(neuralcrest)的不同分化方向有关。通过可视化不同分化方向间基因的互作关系,可以发现神经嵴分化的重要调控因子sox10,以及在不同分化方向中调控细胞命运决定的重要互作基因(图3E~图3G)。除了模块内的信息挖掘之外,研究人员还可视化了空间位置临近的模块之间的基因互作关系。例如,构建了脊索/肋板(notochord/floorplate)相关模块前神经脊椎骨(M13)与临近空间模块肌肉系统(M1)、脊髓(M5)之间互作关系(图3A、图3H),挖掘了在notochord/floorplate中表达的重要分子shha,对周围组织发育的调控作用(图3I、图3J)。
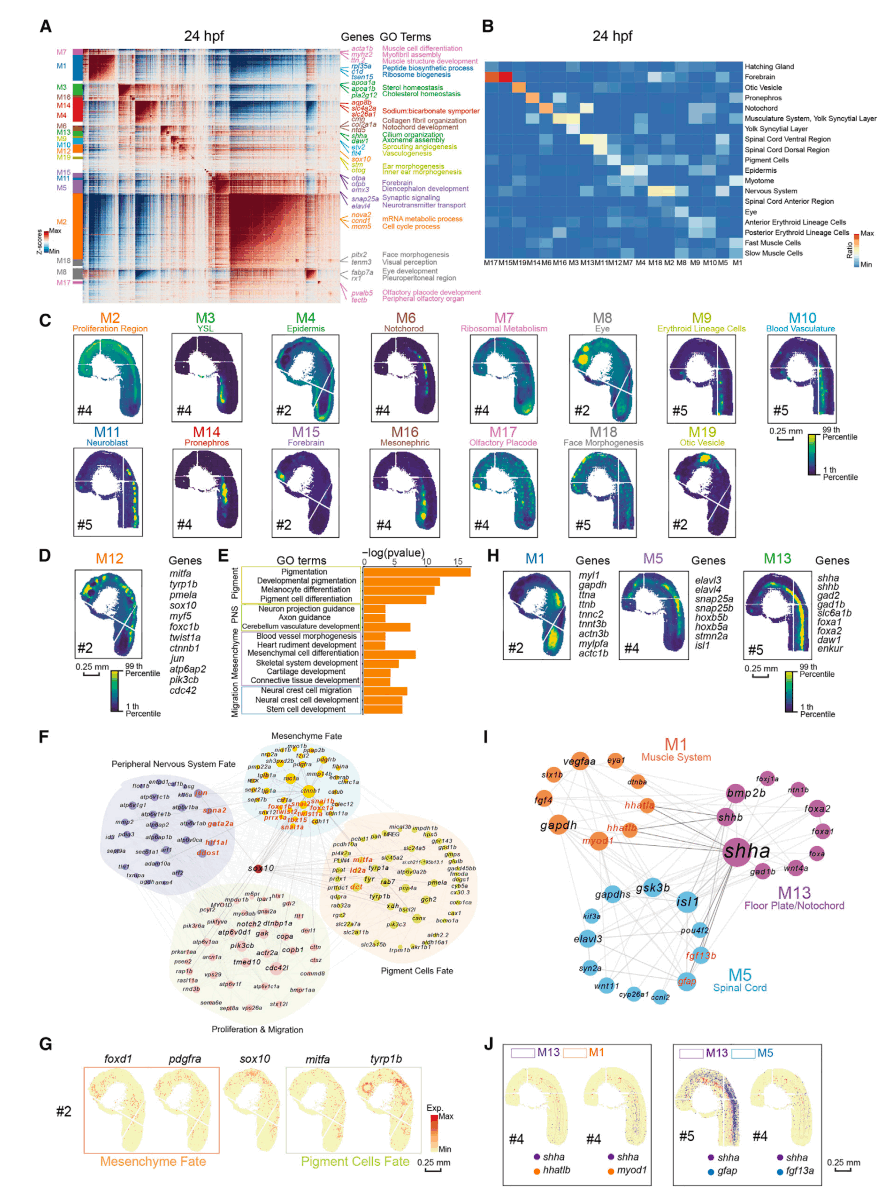
图3 空间共变基因模块揭示不同空间功能区域之间的相互作用关系
4. 整合scRNA-seq和Stereo-seq数据,构建具有空间信息的细胞发育轨迹
为了更精确地描绘胚胎发育过程中的细胞分化轨迹,研究人员选取了与时空转录组6个时间点相对应的斑马鱼胚胎,利用华大自主研发的DNBelab C4平台进行单细胞测序。在完成scRNA-seq数据聚类后,利用SPOTlight整合scRNA-seq和Stereo-seq数据,预测每一个单细胞类群的空间位置(图4A)。例如,3.3 hpf胚胎SPOTlight整合预测的scRNA-seq细胞分群位置结果,与Stereo-seq数据中不同分群的空间位置高度一致,提示了SPOTlight整合分析二者数据的准确性。紧接着,研究人员先利用scRNA-seq数据绘制细胞分化轨迹的桑基图,再将scRNA-seq的不同分化轨迹,利用SPOTlight映射到6个时间点胚胎切片的不同空间位置上,描述了细胞分化过程中的空间分布及动态变化(图4B~图4E)。从桑基图的分支结果来看,来自于同一祖细胞的子细胞,在发育的特定时间点走向不同的细胞命运(图4F)。通过SPOTlight映射,发现在子细胞分化到不同命运之前的上一个时间点,其祖细胞就呈现出不同的空间分布,且此时调控不同命运走向的转录因子与相应祖细胞具有相似的空间分布,从而调控不同祖细胞的命运决定(图4G~图4I)。综上,细胞所处的空间微观环境与其分化方向密切相关,而这一信息仅凭单细胞转录组的数据是无法获得的。

图4 整合scRNA-seq和Stereo-seq数据,构建细胞时空纬度上的发育轨迹
5. 斑马鱼胚胎发育进程中潜在互作配体-受体对的时空分布动态变化
除细胞内因子外,细胞外如配体-受体对的互作,对胚胎发育也起着至关重要的调控作用。研究人员通过计算在斑马鱼胚胎6个发育关键时间点中不同受体-配体对之间的相对距离,推测配体-受体之间的潜在互作关系(图5A)。通过与随机基因间距离的比较,以及空间分布相关性的筛选,设定至少在一个时间点空间相对距离小于bin5的配体-受体对具有潜在互作的可能。通过计算潜在互作配体-受体对的表达强度和相对距离,并根据不同时间点对它们进行排序,探掘不同互作配体-受体对在胚胎发育中的动态变化(图5B)。由于Notch信号通路在胚胎发育中发挥重要作用,研究人员首先通过不同方式展现了Nocth信号通路中的潜在配体-受体对动态变化。随后,计算出不同时间点中与Notch信号通路具有空间相似分布的基因模块(图5C),通过GO分析发现,Nocth信号通路在10、12 hpf两个时间点主要调控体节的形成;18 hpf时,在体节生成和神经系统区域都存在互作;24 hpf时则主要集中在神经系统,表明在斑马鱼胚胎发育不同时间点,Notch信号通路对不同组织器官的发育起到调控作用,这一结论也与前人报道一致。此外,研究人员还通过潜在配体-受体对动态变化,挖掘了一些新的互作模式(图5D~图5H),例如Midkine(mdk)家族基因在胚胎发生中参与神经系统的生成。该研究结果数据系统地揭示了与配体mdka在不同时间段及不同组织结构内发生互作的一系列受体,给后续的调控机制研究提供了参考。

图5 斑马鱼胚胎发育中潜在互作配体-受体对的时空分布动态变化
总结
该研究利用Stereo-seq时空转录组技术,以受精后24h之内6个关键时间点的斑马鱼胚胎为研究对象,绘制了斑马鱼胚胎发育的空间转录图谱。在10μm的分辨率下,对不同时间点的斑马鱼胚胎进行了空间区域的划分,区分出不同组织边界。同时通过进一步的细化分群,在空间位置上区分出空间位置临近的不同细胞亚群和组织。由此研究人员得以在超高分辨率下,得到单细胞测序得不到的信息,如后脑干细胞分化异质性的空间分布差异等。
同时根据空间分布相似性,在不同时间点的斑马鱼胚胎上划分出基因表达共变模块,通过相同模块或不同模块之间基因互作网络分析,进一步探究了空间功能区域间的相互调控作用。该研究选取时空组相对应时间点的胚胎,进行单细胞测序,并利用SPOTlight整合scRNA-seq和Stereo-seq数据,绘制了斑马鱼胚胎的空间发育轨迹。最后,通过计算空间中不同时间点中配体-受体对的相对距离,发现在胚胎发育中具有潜在互作关系的配体-受体对,并确认了其在发育中具有的潜在调控功能。本研究结果将为进一步了解脊椎动物胚胎发育提供理论参考。
作者:刘畅
